の 興奮性シナプス後電位 ニューロンのシナプス後膜における刺激的な可能性です。個々の電位は空間的および時間的に合計され、したがって活動電位を作成できます。重症筋無力症または他の筋無力症などの伝達障害は、これらのプロセスを混乱させます。
興奮性シナプス後電位とは何ですか?

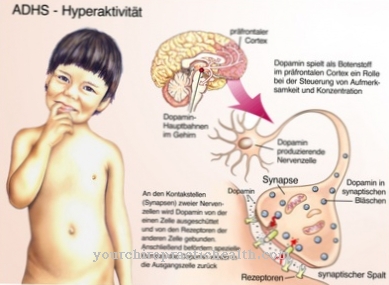
ニューロンは、20〜30 nmのギャップ(シナプスギャップとも呼ばれます)によって互いに分離されています。これは、ニューロンのシナプス前膜領域と下流の神経細胞のシナプス後膜領域の間の最小のギャップです。
ニューロンは興奮を伝達します。したがって、それらのシナプスギャップは、神経伝達物質としても知られている生化学的メッセンジャー物質の放出によって埋められます。これは、下流の細胞の膜領域に興奮性シナプス後電位を作成します。これは、シナプス後膜電位の局所的に限定された変化です。この電位の段階的な変化は、シナプス後部の活動電位を引き起こします。興奮性シナプス後電位はニューロンの興奮伝導の一部であり、下流の細胞膜が脱分極すると発生します。
刺激的なシナプス後電位は、空間的および時間的に加算することにより、次のニューロンによって受信および処理されます。細胞の閾値電位を超えると、新しく形成された活動電位が軸索によって運び去られます。
興奮性シナプス後電位の反対は、抑制性シナプス後電位です。これはシナプス後膜の過分極につながり、活動電位の誘発を防ぎます。
機能とタスク
刺激的なシナプス後電位と抑制性シナプス後電位は、すべての神経細胞に影響を与えます。それらの閾値電位を超えると、神経細胞は脱分極します。彼らは興奮性神経伝達物質を放出することによってこの脱分極に応答します。これらの物質の特定の量は、ニューロンのトランスミッタ感受性イオンチャネルをアクティブにします。これらのチャネルは、カリウムおよびナトリウムイオンを透過します。したがって、興奮性電位という意味での局所電位と段階電位は、ニューロンのシナプス後膜を脱分極させます。
膜電位が細胞内に由来する場合、興奮性シナプス後電位は、体細胞膜の脱分極です。この脱分極は、受動的な伝播の結果として起こります。個々のポテンシャルの合計があります。放出される神経伝達物質の量と一般的な膜電位のサイズにより、興奮性シナプス電位の程度が決まります。膜の脱分極が高ければ高いほど、興奮性シナプス後電位は低くなります。
膜が静止電位を超えてすでに脱分極している場合、シナプス後興奮電位は低下し、特定の状況下ではゼロに達します。この場合、興奮性電位の逆転電位に達します。前脱分極がさらに高いことが判明した場合、反対の符号を持つ電位が発生します。したがって、興奮性シナプス後電位は常に脱分極と同一視される必要はありません。これは、膜を特定の平衡電位に向かって移動させます。平衡電位は、それぞれの静止膜電位よりも下に留まることがよくあります。
これには、複雑なイオン機構の働きが関与しています。興奮性シナプス後電位により、カリウムおよびナトリウムイオンの膜透過性の増加を観察できます。一方、ナトリウムおよびカリウムイオンの導電率が低下した電位も発生する可能性があります。この文脈では、イオンチャネルメカニズムが、すべての漏れやすいカリウムイオンチャネルの閉鎖の引き金であると考えられています。
抑制性シナプス後電位は興奮性シナプス後電位の反対です。ここでも、膜電位は神経細胞のシナプス後膜で局所的に変化します。細胞膜の過分極はシナプスで起こり、興奮性シナプス後電位の枠組み内で活動電位の誘発を阻害します。抑制性シナプスの神経伝達物質は細胞応答を引き起こします。シナプス後膜のチャネルが開き、カリウムまたは塩化物イオンを通過させます。結果として生じるカリウムイオンの流出と塩化物イオンの流入は、シナプス後膜に局所的な過分極を引き起こします。
あなたはここであなたの薬を見つけることができます
muscle筋力低下の薬病気と病気
さまざまな疾患が個々のシナプス間のコミュニケーションを妨害し、したがって化学シナプスでのシグナル伝達も妨害します。 1つの例は、筋終板に影響を与える神経筋疾患重症筋無力症です。これは、これまで原因が不明だった自己免疫疾患です。病気の場合、体は体自身の組織に対する自己抗体を形成します。筋肉疾患では、これらの抗体は神経筋終板のシナプス後膜に向けられています。ほとんどの場合、この疾患の自己抗体はアセチルコリン受容体抗体です。それらは神経と筋肉の間の接続点でニコチン性アセチルコリン受容体を攻撃します。結果として生じる免疫学的炎症は、局所組織を破壊します。
その結果、アセチルコリンとその受容体との間の相互作用がアセチルコリン受容体抗体によって困難にされたり、阻止されたりするため、神経と筋肉の間のコミュニケーションが妨げられます。したがって、活動電位はもはや神経から筋肉に伝わることができません。したがって、筋肉は興奮しなくなります。
すべてのアセチルコリン受容体の合計は、受容体が免疫活動によって破壊されると同時に減少します。シナプス膜は崩壊し、エンドサイトーシスはオートファゴソームを作成します。輸送小胞はオートファゴソームと融合し、この免疫反応の結果としてアセチルコリン受容体が変化します。これらの変更により、モーターのエンドプレート全体が変更されます。シナプスのギャップが広がります。このため、アセチルコリンはシナプス間隙から拡散するか、受容体に結合せずに加水分解されます。
他の筋無力症は、シナプス裂と興奮性シナプス後電位に同様の効果を示します。



.jpg)