核酸塩基 DNAおよびRNA分子の長鎖がリン酸化ヌクレオチドの形で構成されるビルディングブロックです。
ロープに似た二本鎖を形成するDNAでは、4つの存在する核酸塩基が水素ブリッジを介してそれぞれの相補的な塩基と固体のペアリングを形成します。核酸塩基は、二環式プリンまたは単環式ピリミジン骨格のいずれかで構成されます。
核酸塩基とは何ですか?
長いDNA二重らせん分子鎖の構成要素としての4つの核酸塩基アデニン、グアニン、シトシン、チミンは、アデニン-チミン(A-T)とグアニン-シトシン(G-C)の一定のペアリングを形成します。
アデニンとグアニンの2つの塩基はそれぞれ、プリン基本構造の修飾された二環式の6環と5環で構成されているため、プリン塩基とも呼ばれます。他の2つの核酸塩基であるシトシンとチミンの基本構造は、修飾ピリミジン骨格に対応する複素環式芳香族6員環で構成されているため、ピリミジン塩基とも呼ばれます。 RNAは主に一本鎖として存在するため、最初は塩基対はありません。これは、mRNA(メッセンジャーRNA)を介した複製中にのみ行われます。
RNA鎖のコピーは、DNAの2番目の鎖に類似した相補的核酸塩基で構成されています。唯一の違いは、RNAではチミンの代わりにウラシルが使用されていることです。 DNAおよびRNA鎖分子は、核酸塩基によって純粋な形で形成されるのではなく、代わりに、DNAの場合は、5-糖デオキシリボースと結合して対応するヌクレオシドを形成します。 RNAの場合、糖グループはリボースで構成されます。さらに、ヌクレオシドはリン酸残基でリン酸化され、いわゆるヌクレオチドを形成します。
プリン塩基のヒポキサンチンとキサンチンは、DNAとRNAにも含まれ、修飾されたチミンに対応します。ヒポキサンチンは、アミノ基(-NH3)をヒドロキシル基(-OH)で置き換えることによりアデニンから形成され、キサンチンはグアニンから形成されます。両方の核酸塩基は遺伝情報の伝達に寄与しません。
機能、効果、タスク
DNAの二本鎖を構成する核酸塩基の最も重要な機能の1つは、目的の位置に存在を示すことです。
核酸塩基の配列は遺伝暗号に対応し、タンパク質を構成するアミノ酸の種類と配列を定義します。これは、DNAの構成要素としての核酸塩基の最も重要な機能は、受動的で静的な役割で構成されていることを意味します。つまり、それらは代謝に積極的に介入せず、読み取りプロセス中にメッセンジャーRNA(mRNA)によって生化学的構造が変化することはありません。それはDNAの寿命を部分的に説明します。
ミトコンドリアDNA(mtDNA)の半減期は、核酸塩基間に元々存在していた結合の半分が崩壊する間に、環境条件に大きく依存し、正の温度の平均的な条件下では約520年から永久凍土条件下では最大150,000年まで変化します。 。
RNAの一部として、核酸塩基はやや積極的な役割を果たします。原則として、細胞が分裂すると、相補的な鎖を形成できるようにするためにDNA二本鎖が分解され、互いに分離されます目的のタンパク質が組み立てられます。別の核酸塩基であるジヒドロウラシルは、タンパク質合成中にアミノ酸を輸送するために使用される、いわゆる輸送RNA(tRNA)でのみ発生します。
一部の核酸塩基は、特定の生化学的プロセスを積極的に触媒的に可能にし、制御する酵素の一部として、完全に異なる機能を果たします。アデニンは、細胞のエネルギーバランスにおけるヌクレオチドとして最もよく知られているタスクを実行します。アデニンは、アデノシン二リン酸(ADP)およびアデノシン三リン酸(ATP)とニコチンアミドアデニンジヌクレオチド(NAD)のコンポーネントとして電子ドナーとして重要な役割を果たします。
教育、発生、特性および最適値
非リン酸化型では、核酸塩基は炭素、水素、酸素のみからなり、ユビキタスで自由に利用できる物質です。したがって、体は核酸塩基自体を合成することができますが、そのプロセスは複雑でエネルギーを消費します。
したがって、リサイクルによる核酸の回収が好ましい。 B.特定の化合物を含むタンパク質を分解し、エネルギーをほとんど消費せずに、またはエネルギーを獲得するだけで、核酸に分離および変換できる。核酸は通常、体内で純粋な形では発生しませんが、リボースまたはデオキシリボース分子が付着したヌクレオシドまたはデオキシヌクレオシドとしてほとんど発生します。 DNAおよびRNAの構成要素として、また特定の酵素の構成要素として、核酸またはそれらのヌクレオシドは、1〜3個のリン酸基(PO4-)で可逆的にリン酸化されます。
核酸塩基の最適な供給に関する基準値はありません。核酸塩基の欠乏または過剰は、代謝の特定の障害を介して間接的にのみ決定できます。
病気と障害
核酸塩基に関連してもたらされる危険、妨害、およびリスクの性質は、DNAまたはRNA鎖の数と配列のエラーであり、タンパク質合成のコーディングに変化をもたらします。
身体がその修復メカニズムを介して障害を治療できない場合、それは生物学的に不活性または使用可能なタンパク質の合成に至り、次に軽度から重度の代謝障害につながる可能性があります。たとえば、 B.遺伝子変異が存在し、これは、代謝障害を介して最初から症候性疾患を引き起こす可能性があり、これは不治の可能性がある。しかし、健康なゲノムであっても、DNAおよびRNA鎖の複製中にコピーエラーが発生し、代謝に影響を与える可能性があります。
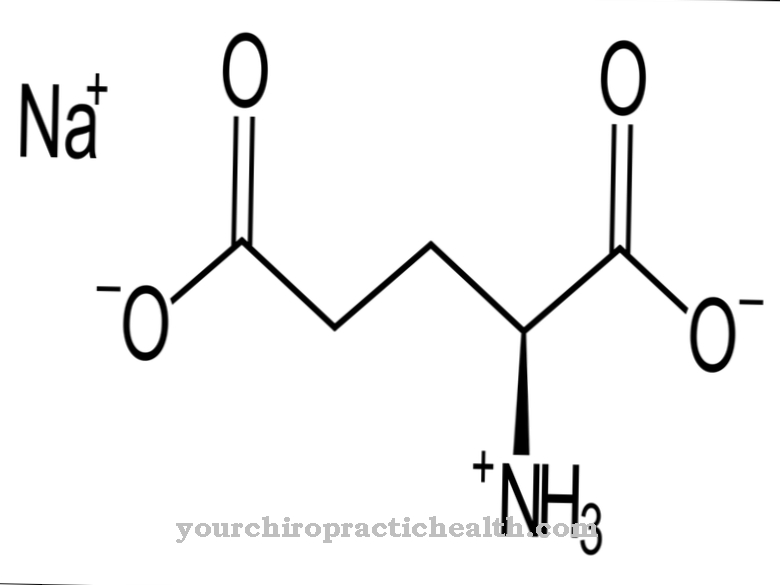
プリンバランスの既知の代謝障害はzです。 B. x染色体の遺伝的欠陥に戻る。遺伝的欠陥のため、プリンベースのヒポキサンチンとグアニンはリサイクルできず、最終的には関節の尿路結石と痛風の形成を促進します。














.jpg)