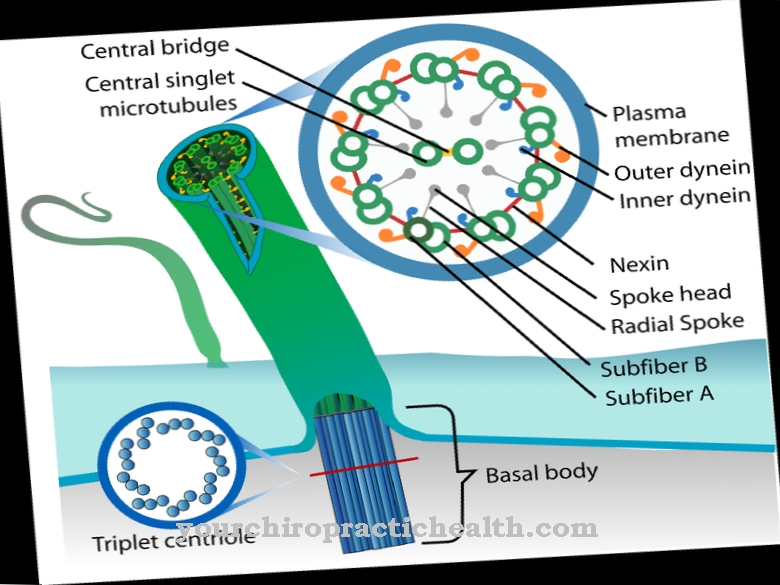
の スプライス 真核生物の細胞核における転写中の重要なプロセスを表しており、その間に成熟mRNAがプレmRNAから出現します。転写後もプレmRNAにまだ含まれているイントロンが削除され、残りのエクソンが結合されて、完成したmRNAが形成されます。
スプライシングとは

分子生物学の中心的な教義では、遺伝情報の流れは、情報担体DNAからRNAを介してタンパク質に至るとされています。遺伝子発現の最初のステップは、転写と呼ばれるものです。 RNAはDNAをテンプレートとして使用して合成されます。 DNAは遺伝情報の担体であり、アデン、チミン、グアニン、シトシンの4つの塩基からなるコードを使用してそこに保存されます。 RNAポリメラーゼタンパク質複合体は、転写中にDNAの塩基配列を読み取り、対応する「プレメッセンジャーRNA」(略してプレmRNA)を生成します。チミンの代わりに、ウラシルは常に組み込まれています。
遺伝子はエクソンとイントロンで構成されています。エキソンは、実際に遺伝情報をコードするゲノムの一部です。対照的に、イントロンは遺伝子内の非コードセクションを表します。DNAに格納された遺伝子は、後のタンパク質のアミノ酸に対応せず、翻訳に寄与しない長いセクションが横断します。
遺伝子には最大60個のイントロンがあり、長さは35〜100,000ヌクレオチドです。平均して、これらのイントロンはエクソンよりも10倍長い。転写の最初のステップで生成されたpre-mRNA(未成熟なmRNAとも呼ばれます)には、依然としてエクソンとイントロンの両方が含まれています。これが接合プロセスの始まりです。
イントロンはプレmRNAから削除する必要があり、残りのエクソンは一緒にリンクする必要があります。その後、成熟したmRNAは細胞核を離れて翻訳を開始できます。
スプライシングは主にスプライセオソーム(ドイツ語:スプライセオソーム)を利用して行われます。これは5つのsnRNP(小さな核リボ核タンパク質粒子)で構成されています。これらの各snRNPは、snRNAとタンパク質で構成されています。 snRNPの一部ではない他のいくつかのタンパク質も、スプライセオソームの一部です。スプライソソームは、メジャーとマイナーのスプライソソームに分けられます。主要なスプライセオソームはすべてのヒトイントロンの95%以上を処理し、マイナーなスプライセオソームは主にATACイントロンを処理します。
スプライシングの説明として、リチャードジョンロバーツとフィリップA.シャープが1993年にノーベル医学賞を受賞しました。 Thomas R. CechとSidney Altmanは、RNAの選択的スプライシングと触媒作用に関する研究で1989年にノーベル化学賞を受賞しました。
機能とタスク
スプライシングプロセス中に、スプライセオソームは個々の部分から新しく形成されます。哺乳類では、snRNP U1は最初に5’-スプライス部位に付着し、残りのスプライセオソームの形成を開始します。 snRNP U2はイントロンの分岐点に結合します。その後すぐに、tri-snRNPもバインドします。
スプライセオソームは、2つの連続したエステル交換反応によってスプライシング反応を触媒します。反応の最初の部分では、「分岐点シーケンス」(BPS)のアデノシンの2'-OH基の酸素原子が、5 'スプライス部位のホスホジエステル結合のリン原子を攻撃します。これにより、5 ’エクソンが放出され、イントロンが循環します。 5'-エクソンの現在フリーの3'-OHグループの酸素原子は、3'-スプライス部位に結合します。その結果、2つのエクソンが接続され、イントロンが解放されます。イントロンは、ラリアットと呼ばれる合理化された構造になり、その後分解されます。
これとは対照的に、スプライソソームは自己スプライシングに関与しません。ここでは、イントロンはRNA自体の二次構造によって翻訳から除外されています。 tRNA(トランスファーRNA)の酵素によるスプライシングは真核生物と古細菌で発生しますが、細菌では発生しません。
スプライシングプロセスは、エクソンとイントロンの境界で正確に行われる必要があります。これは、1つのヌクレオチドだけがずれると、アミノ酸が正しくコーディングされず、完全に異なるタンパク質が形成されるためです。
pre-mRNAのスプライシングは、環境の影響や組織の種類によって異なる結果になる場合があります。つまり、同じDNA配列から、したがって同じプレmRNAから、異なるタンパク質を形成することができます。このプロセスは、代替スプライシングとして知られています。人間の細胞には約20,000の遺伝子が含まれていますが、選択的スプライシングにより数十万のタンパク質を生成することができます。すべての人間の遺伝子の約30%に選択的スプライシングがあります。
スプライシングは進化において主要な役割を果たしてきました。多くの場合、エクソンはタンパク質の個々のドメインをコード化しており、さまざまな方法で互いに組み合わせることができます。これは、完全に異なる機能を持つ多種多様なタンパク質がほんの数個のエクソンから生成できることを意味します。このプロセスはエクソンシャッフリングと呼ばれます。
病気と病気
一部の遺伝性疾患はスプライシングと密接に関連している可能性があります。非コード化イントロンの変異は通常、タンパク質の形成にエラーを引き起こしません。ただし、スプライシングの調節に重要なイントロンの一部に変異が発生した場合、pre-mRNAのスプライシングに障害が発生する可能性があります。結果として得られる成熟したmRNAは、不完全な、または最悪の場合は有害なタンパク質をエンコードします。これは、たとえば、ある種のベータサラセミア、遺伝性貧血の場合です。このようにして発症する疾患の他の代表例は、例えば、エーラース・ダンロス症候群(EDS)II型および脊髄性筋萎縮症です。


.jpg)