アデニン 有機核塩基として、他の3つの塩基と一緒に、DNAおよびRNAの遺伝情報の基本的なビルディングブロックの1つを形成する、プリン骨格を持つヘテロ二環式芳香族化合物です。
さらに、ヌクレオシドまたはヌクレオチドの形のアデニンは、NAD、FADH2、またはATPとしての代謝、特にミトコンドリアの細胞のエネルギーバランスにおいて重要な役割を果たします。
アデニンとは
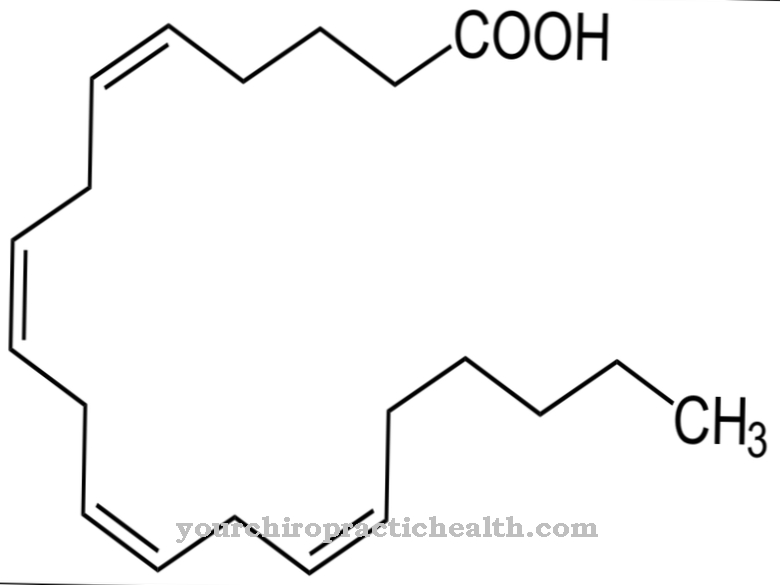
化学分子式を持つアデニン C5N5H5 アミノ基(NH2)が結合したヘテロ二環式芳香環(プリン基本構造)で構成されます。したがって、アデニンは アミノプリン 専用。これは、摂氏220度で昇華する淡黄色の固体です。つまり、直接気体状態に変化し、水にほとんど溶けません。
デオキシリボース糖分子を追加すると、アデニンがデオキシアデノシンに変わります。これは、二重らせんDNAを構成する4つのビルディングブロックの1つです。チミジンと付加されたデオキシリボース分子から形成されるデオキシチミジンは、相補的な塩基として機能します。 RNAの場合、これは少し変更されたプロセスです。アデニンは、D-リボース糖分子の付加によってアデノシンになります。アデノシンは、RNAのDNAにおけるデオキシアデノシンの位置を占めます。相補的な塩基はチミンではなく、ウリジンの形のウラシルです。
さらに、アデノシンはヌクレオチドATP、ADPおよびAMPの基本構造を形成し、細胞のエネルギーバランスに重要な役割を果たします。アデノシンはまた、補酵素A、NADPH、NADHなどの多くの酵素、ホルモン、神経調節因子の補因子として重要な機能を果たします。
機能、効果、タスク
DNA二重らせんの鎖の一部として、アデノシンは、デオキシチミジンの形で相補的な核酸塩基チミンと2つの水素ブリッジを介してアデニン-チミン塩基対(A-T)を形成します。ほとんどが一本鎖のRNAでは、アデニンは類似の機能を持っていますが、相補鎖であるmRNA(メッセンジャーRNA)の形成では、相補的な塩基はチミジンではなくウラシルです。
DNAおよびRNAの成分として、アデニンは代謝プロセスに直接関与していませんが、他の核酸塩基と組み合わせて対応するタンパク質を合成するためのアミノ酸配列をコード化するためにのみ機能します。呼吸鎖と呼ばれるほとんどすべての細胞のエネルギー代謝の一部は、本質的に一連の酸化および還元プロセス、いわゆるレドックスプロセスで構成されています。アデノシンはリン酸化されてアデノシン三リン酸(ATP)を形成しますが、呼吸鎖の中心にあります。 ATPはリン基を放出し、したがってアデノシン二リン酸(ADP)またはアデノシン一リン酸(AMP)になります。全体として、それは炭水化物の分解を使用して代謝のためのエネルギーを生成する発熱プロセスです。 B.筋肉の働きに備える。
この機能では、アデニンまたはアデノシンが直接化学反応に関与しています。酸化還元反応の連鎖における重要な動的成分は、水素(H)または他の電子キャリアに結合した電子からの電子移動でもあります。ここでも、アデニンとアデノシンはニコチンジアミド(NAD)やその他の酵素や触媒の機能的成分であり、最終的には水素の水への酸化(燃焼)を多くの触媒的に制御された個々のステップに分解するため、熱傷による損傷を引き起こすことなく代謝に利用できます。
教育、発生、特性および最適値
化学式C5N5H5によると、アデニンは基本的なビルディングブロックの炭素、窒素、水素で構成されており、それらはすべて自然界に豊富にあります。希少な微量元素やミネラルは必要ありません。したがって、合成のための原材料の不足を恐れる必要はなく、身体自体の生産プロセスの問題です。
合成には時間がかかり、エネルギーを大量に消費するため、身体が別の経路を使用する時間の約90%は、リサイクルによってアデニンを合成します。プリン代謝の過程で、アデニンはより複雑な化合物の分解生成物として得られます。アデニンは、デオキシリボースの分子の追加によってのみヌクレオシドとして生化学的に有効になります。これはアデニンをデオキシアデノシンに変えます。さらに1〜3個のリン酸残基を追加すると、デオキシアデノシンは、アデノシン一リン酸(AMP)、環状アデノシン一リン酸(cAMP)、アデノシン二リン酸(ADP)、またはアデノシン三リン酸(ATP)と呼ばれるヌクレオチドになります。
動的に変化する要件を満たし、体の循環で遊離アデニンが発生しない環境でのアデニンの多様なタスクとその生化学的活性発現に従って、アデニンレベルは測定できません。無傷のプリン代謝に関する間接的な結論は、特定の代謝プロセスを観察および測定することによってのみ導き出すことができます。
病気と障害
最もよく知られている-デファクトですがまれな-身体自身のアデニンの生産とその生理活性型に関連して発生する代謝障害は、レッシュナイハン症候群です。これは、x染色体の遺伝的欠陥です。遺伝子変異はヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRT)の完全な欠乏につながります。
HGPRTの欠如は、プリン代謝の障害につながるため、通常行われるプリン塩基のヒポキサンチンとグアニンのリサイクルはもはや必要ありません。代わりに、体は新合成によってアデニンを継続的に生成することを余儀なくされます。これは、過剰な量の尿酸と尿酸結晶の沈殿を引き起こし、痛風や関節の尿路結石の形成を引き起こす可能性があります。さらに、新生児は通常、精神発達障害と自己攻撃の増加を経験します。
もう一つのまれな遺伝性疾患はハンチントン病です。 4番染色体に遺伝的欠陥があります。通常、シトシン-アデニン-グアニンの塩基配列は10〜30回の繰り返しで特定の遺伝子に見られます。遺伝子変異に起因するこれらのいわゆるトリプレット反復が36を超えると、遺伝性疾患のハンチントン病が発生します。病気の過程で、治癒できない運動障害と神経損傷があります。