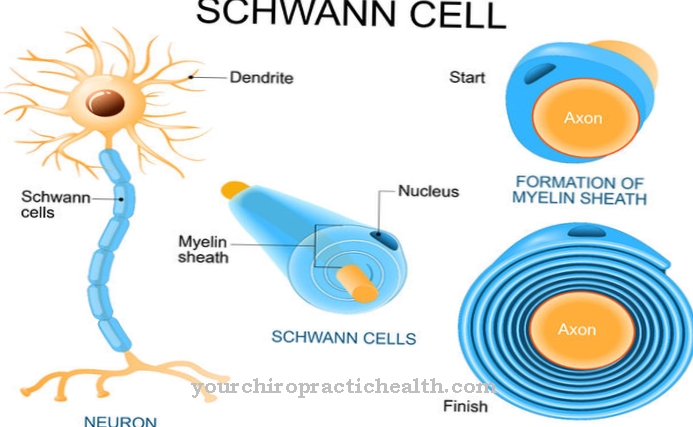
ヒストン 細胞核の一部です。それらの存在は、単細胞生物(細菌)と多細胞生物(人間、動物、または植物)の際立った特徴です。ヒストンに類似したタンパク質を持っている細菌株はほとんどありません。進化は、高等生物の細胞内に遺伝物質としても知られている非常に長いDNA鎖をより良く、より効果的に収容するためにヒストンを生成しました。なぜなら、ヒトゲノムがばらばらになっている場合、細胞が存在する細胞の段階に応じて、1〜2 m程度の長さになるからです。
ヒストンとは何ですか?
より高度に発達した生物では、ヒストンは細胞核で発生し、正に帯電したアミノ酸(特にリジンとアルギニン)の割合が高くなります。ヒストンタンパク質は、H1、H2A、H2B、H3、およびH4の5つの主要なグループに分類されます。 H2A、H2B、H3、およびH4の4つのグループのアミノ酸配列は、異なる生物の間でほとんど違いがありませんが、H1、接続ヒストンには、より多くの違いがあります。 核を含む鳥の赤血球の場合、H1はH5と呼ばれる別の主要なヒストングループで完全に置き換えられています。
ほとんどのヒストンタンパク質の配列の大きな類似性は、ほとんどの生物でDNAの「パッケージング」が同じ方法で行われ、結果として得られる3次元構造がヒストンの機能に等しく有効であることを意味します。進化の過程で、ヒストンの発達は非常に早く起こったに違いなく、哺乳類や人間が出現する前でも維持されていたに違いありません。
解剖学と構造
細胞内の個々の塩基(ヌクレオチドと呼ばれる)から新しいDNA鎖が形成されるとすぐに、それを「パッケージ化」する必要があります。この目的のために、ヒストンタンパク質は二量体化し、次にそれぞれが2つの四量体を形成します。最後に、ヒストンコアは、ヒストンオクタマーという2つの四量体で構成されています。ヒストンオクタマーは、DNA鎖が周りを包み込み、部分的に貫通しています。したがって、ヒストンオクタマーは、ねじれたDNA鎖内の3次元構造に配置されます。
DNAが周囲にある8つのヒストンタンパク質は、ヌクレオソームの複合体全体を形成します。 2つのヌクレオソーム間のDNAの領域はリンカーDNAと呼ばれ、約20〜80ヌクレオチドを含みます。リンカーDNAは、ヒストンオクタマーの「進入」と「離脱」に関与します。したがって、ヌクレオソームは約146ヌクレオチド、リンカーDNA部分、および8つのヒストンタンパク質で構成されているので、146ヌクレオチドはヒストンオクタマーの周りを1.65回ラップします。
さらに、各ヌクレオソームはH1分子に関連付けられているため、DNAの入口と出口はヒストンによって結合され、DNAのコンパクトさが増します。ヌクレオソームの直径は約10〜30 nmで、多くのヌクレオソームは、電子顕微鏡で見ると真珠のひものように見える長いDNAヒストン鎖であるクロマチンを形成しています。ヌクレオソームは、ひも状のDNAに囲まれた、または接続された「真珠」です。
いくつかの非ヒストンタンパク質は、個々のヌクレオソームの形成、またはクロマチン全体の形成をサポートし、細胞が分裂する場合、最終的に個々の染色体を形成します。染色体はクロマチンの最大タイプの圧縮であり、細胞の核分裂中に光学顕微鏡で認識できます。
機能とタスク
上記のように、ヒストンは正電荷を持つ基本的なタンパク質であるため、静電引力によって負に帯電したDNAと相互作用します。 DNAはヒストンオクタマーを「包み込む」ため、DNAはよりコンパクトになり、各細胞の核にフィットします。 H1は、高レベルのクロマチン構造を圧縮する機能を備えており、ほとんどの場合、転写、つまり翻訳、つまりmRNAを介したこのDNA部分のタンパク質への翻訳を防止します。
細胞が「休止」している(間期)か、分裂しているかに応じて、クロマチンは多少濃縮または濃縮されています。間期では、クロマチンの大部分はあまり凝縮されていないため、mRNAに転写されます。つまり、読み取られ、後でタンパク質に翻訳されます。ヒストンは、その近傍にある個々の遺伝子の遺伝子活性を調節し、mRNA鎖の転写と作成を可能にします。
細胞が分裂し始めると、DNAはタンパク質に翻訳されず、作成された2つの娘細胞に均等に分配されます。したがって、クロマチンはヒストンによって強く凝縮され、さらに安定化されます。染色体が見えるようになり、他の多くの非ヒストンタンパク質の助けを借りて、新しく出現した細胞に分配することができます。
病気
ヒストンは、新しい生物の創造に不可欠です。ヒストン遺伝子の変異により、1つ以上のヒストンタンパク質を形成できない場合、この生物は生存できず、その後の発生は時期尚早に停止されます。これは主にヒストンの高度な配列保存によるものです。
しかし、さまざまな悪性脳腫瘍のある子供や成人では、腫瘍細胞のさまざまなヒストン遺伝子に変異が生じる可能性があることが以前から知られています。ヒストン遺伝子の変異は、いわゆる神経膠腫で上記に説明されています。細長い染色体の尾もこれらの腫瘍で発見されています。テロメアと呼ばれる染色体のこれらの末端部分は、通常、染色体の寿命に関与しています。この文脈において、ヒストン変異を有する腫瘍における伸長したテロメアは、これらの変性した細胞に生存上の利点を与えると思われる。
一方で、さまざまなヒストン遺伝子に変異があり、そのために規制タスクを実行しない、または不十分にしか実行しない変異ヒストンタンパク質を産生する他のタイプの癌が知られています。これらの発見は現在、特に悪性で攻撃的な腫瘍に対する治療法の開発に使用されています。

.jpg)